By
ADRIAN J. SHINE
Loch
Ness and Morar Project
DAVID S. MARTIN
Loch
Ness and Morar Project
ROSALIND S. MARJORAM
Loch Ness and Morar Project
Introduction
The first suggestions
of an off-shore population of Charr Salvelinus
alpinus in Loch Ness were made as a result
of echo-sounding by Dr. P.F. Baker (Baker and
Westwood, 1960). Echo-sounder transects were also made as
a part of the Institute of Terrestrial Ecology's
multi-disciplinary survey during 1977-80 (Maitland,
1981).
Throughout the 1980s,
the Loch Ness and Morar Project conducted a pelagic
programme of qualitative echo-sounding, gill-netting
and trawling, designed to reveal the basic distributions
of fish within the water column and along the
axis of the loch (Shine and Martin, 1988). Species taken by pelagic gill-netting and
trawling consist mainly of Charr, with some Brown
Trout Salmo
trutta and, particularly in the southern basin, Three-spined Sticklebacks Gasterosteus aculeatus. From
1988 onwards, more quantitative acoustic methods
have been applied, which have improved our understanding
of the distribution, numbers, biomass, and diurnal
behaviour of the fish population. Observations
have also been made of the vertical and horizontal
distribution of zooplankton.
Most of these exercises
were carried out as part of 'Operation Echo',
a series of collaborations with the Simrad Company
and the Marine Laboratory of the Department of
Agriculture and Fisheries (D.A.F.S.) at Aberdeen. Simultaneous surveys were also carried
out by the Hydroacoustic Unit from Royal Holloway
University of London, and are described separately
(Kubecka, Duncan and Butterworth, 1993).
Vol 105, The Scottish
Naturalist: Pelagic Fish and Zooplankton in Loch
Ness p197
Methods
The data presented here
has been obtained from several sources: from a
fixed station established on deep-water moorings,
and from various longitudinal and transverse runs
with a variety of vessels (Figure
1, 9K).
Gill nets, ranging in
mesh size (stretched diagonal) from 30 to 90 mm,
have been deployed from the fixed station to depths
of over 30 m.
Most trawling was carried
out in 1988 by the D.A.F.S. trawler Goldseeker, using a 'sprat'
trawl with a 12 mm codend. The trawl was towed at 20-30 m depth at
2.0 - 2.5 knots with a vertical mouth opening
of approximately 6.0 m. Depths were established by a Simrad Trawlink
acoustic system. There were four hauls, totalling four hours of trawling.
In November 1992, material
was made available from a large trawl undergoing
configuration trials by D.A.F.S. aboard the research
vessel Calanus.
Plankton observations
are based for the most part upon vertical hauls
at 6.0 m increments using a closing plankton net
with a 28 cm diameter, one metre in length with
a 0.25 mm mesh size. Hauls were taken around mid-day and midnight. Numbers are expressed per cubic metre,
with no allowance for any net factor. The hauls presented for 1983 were taken with a 31-litre Patalas
water sampler.
Longitudinal results are
from the Royal Holloway Clarke-Bumpus sampler
(15 cm aperture) towed at 20 m depth for distances
of one nautical mile, separated by intervals of
one nautical mile.
A Focal Industries Optical
Plankton Counter, model OPC-IT, was used in 1990
to investigate particles within the scattering
layer and water column. This had a sampling aperture of 3.0 x 22 cm
and a 640 nanometer light source with beam dimensions
of 0.4 x 2.0 cm.
Particles were detectable
from 0.25 mm to 2.0 cm, and counts were made by
towing over one minute intervals at depth increments
through layers detected acoustically. A D.A.F.S. high-speed plankton net was attached
to the towed body, and depths were measured using
a Simrad H.P.R. 310T transponder.
Temperature structure
was measured by a variety of instruments, ranging
from a Windermere Profiler to a Lowrance single
thermistor on a depth-marked wire.
Vol 105, The Scottish
Naturalist: Pelagic Fish and Zooplankton in Loch
Ness p198
Acoustic Methods
Most fixed station observations
were made using conventional chart-recording single
beam echo-sounders. A Lowrance X-16 (50 kHz) was used from
the fixed station in the North Basin in 1991 and
1992. This has the facility to reduce paper speed,
to the extent that one hour could be represented
by 2.0 cm of chart, thus permitting monitoring
over several days to be presented in a manageable
form.
Much of the basic distribution
of targets within the water column may be overviewed
in this way. There is, however, a difficulty in quantifying
the numbers of fish and their sizes. Even echo-sounders capable of target strength measurement with a
single beam cannot quantify the true target strength,
since a small fish in the centre of the beam may
appear stronger than a large fish off axis. Acoustic surveys using echo-integration therefore
required scaling factors derived from fish catches.
In the 1980s three new
acoustic methods capable of in
situ target strength assessment were refined,
and between 1988 and 1992 the Project was fortunate
to participate in the first practical use of these
systems in Britain's freshwaters.
All
three systems exploit sophisticated software processing,
in particular to apply 'single fish echo criteria'. This criteria rejects echoes below a given
noise threshold and those of short length likely
to be noise. In addition, echoes larger than the criteria
at various points along the peak are classified
as multiple targets. All systems utilise echo-integrators to
total the acoustic biomass, while relating the
measured target strength distribution to this. The differences lie in the way this target
strength distribution is obtained; detailed descriptions
and comparisons of the three systems are given
below:
1. H.A.D.A.S. Hydroacoustic Data Acquisition System.
See Craig and Forbes (1969). Figure
2a.(4K)
This is an indirect statistical
method for use with a single beam echo-sounder. Developed by Dr.Torfin Lindem of Oslo
University, the software package utilises the
algorithm proposed by Craig and Forbes (1969)
to remove the effect of the beam pattern from
the received echoes.
In general, the echoes are divided into classes, with the strongest assumed
to come from the largest fish in the centre of
the beam, with the second strongest coming from
the second largest fish in the centre, together
with the largest fish in the first off-centre
classification.
Vol 105, The Scottish Naturalist: Pelagic
Fish and Zooplankton in Loch Ness p200
This process is applied progressively throughout the various size classes,
and therefore requires a large number of echoes
(1,000+) and a good percentage of targets passing
the single fish criteria. The proportions between resolved single fish
and multiples are established through echo-integration,
thus allowing the total density to be calculated. The equipment was originally developed
for the Simrad EY-M sounder, and has been used
in a number of surveys (Jurvelius, Lindem and
Louhimo, 1984; Hartmann et
al., 1987).
On 24th July 1988 Dr. Lindem brought an advanced version of H.A.D.A.S. to
Loch Ness, where it was linked to a Simrad EY200
sounder (49 kHz) calibrated with a copper sphere. The equipment was mounted with an in-hull
transducer aboard the Simrad vessel Simson Echo,
and two runs of approximately 5.0 km were made
in the South Basin between Fort Augustus and Invermoriston.
The first run was by day at 15.00 hrs and the second at dusk (20.00 hrs). Single fish resolution was mostly 90-100%
and never below 70%. By day 4,790 single fish echoes were resolved, and by night 1,600. Trawling was carried out simultaneously
in the same area by the Goldseeker.
2.The
Dual Beam Method (Biosonics Inc.).
See Ehrenberg (1978); Traynor and Ehrenbrg (1979). Figure
2b (4K)
This is a direct acoustic method of target strength determination, transmitting
pulses on a narrow beam element and receiving
them on both this transducer and through a wider
surrounding beam. The ratio between the signals received by the
two beams provides the off-axis angle of the target. The Royal Holloway Hydroacoustic Unit brought
a Biosonics 105 Dual Beam sounder to the loch
in May 1991, October 1991 and May 1992. During the May visits the equipment was
used simultaneously with the EK500 Split Beam
equipment described below. The Dual Beam findings are described in
an accompanying paper by Kubecka et al. (1993).
3. The Split Beam Method (Simrad EK500).
See Bodholt, Nes and Solli (1988); Ehrenberg (1979). Figure
2c (4K)
This is another direct method of in situ target strength assessment, and relies upon the phase difference
of echoes received by separate elements of the
transducer, thus locating the target within the
beam and compensating accordingly.
Vol 105, The Scottish Naturalist: Pelagic
Fish and Zooplankton in Loch Ness p201
The Simrad EK500 utilises four transducer elements, and was first used at
Loch Ness with a hull-mounted transducer aboard
the Simson Echo in July 1990. Subsequently
it was installed with both 38 kHz and 120 kHz
transducers rigged at 1.0 m depth aboard the Project
vessel Ecos in May 1991
and 1992.
In May 1992 a series of 0.5 nautical mile runs, totalling 10.5 n-miles,
were made in the North Basin both by day and at
approximately midnight. The distributions quoted are the mean of
these runs.
On 24th May a length run was made along the axis of the loch. The EK500 results are produced through
a colour printer which tabulates target strength
distributions within chosen depth layers. The number of accepted single fish echoes
is also recorded, together with the SA (area back-scattering
coefficient) or integrated value expressed in
m2 per n-miles2.
In the estimation of fish densities the sigmas (linear
values of back-scattering cross-section for individual
fish) of targets in the logarithmic dB scale are
calculated thus:
ó= 4pi (10. TS/10)
Where TS is target
strength.
The percentage
of sigmas per target strength group is then divided
into the SA to give fish per n-mile2 and converted into fish/ha.
Where fish lengths
are suggested they are based upon the formulae
of Lindem (1984):
TS = 20 Log L-68 for small fish
TS = 20 Log L-67 for larger fish
Where L is fish
length in cm.
On occasion, Love's
(1977) empirical formula is also used:
TS = 18.4 Log
L -1.6 Log
F -61.6
Where L is fish
length in cm and F is acoustic frequency in kHz.
Vol
105, The Scottish Naturalist: Pelagic Fish
and Zooplankton in Loch Ness p202
Results
Fish
Figure
3 (13K) provides a backdrop to the summer
vertical and diurnal distributions. The record was taken from a fixed mooring
in the North Basin over a 24-hour period, and
shows individual fish targets ranging to 30 m
throughout the day. A well defined 'scattering layer' makes regular
migrations nearly to the surface at midnight. The layer appears particularly dense at depth within the 33oecho-sounder
beam, but as it approaches the surface in the
narrower part of the beam it can be seen to be
composed of targets which are detected as individuals. The larger fish do not migrate to the surface
at night.
System Comparisons
The
H.A.D.A.S. and EK500 results quantify this process. A comparison between the peak target strengths
within the scattering layer shows the H.A.D.A.S.
(49 kHz) at -54 dB, the EK500 Split Beam (38 kHz)
at -63 dB, and the EK500 Split Beam (120 kHz)
at -70 dB (Figure
4, 16K). The Biosonics Dual Beam (420 kHz) peak
was at -75 dB (Kubecka et al., 1993).
Of these results,
the -54 dB and -63 dB are reasonably compatible
with fish between 4.0 cm and 2.0 cm respectively. The frequency of other targets within the distribution
falls fairly steadily with increasing strength,
and shows less variation between the systems.
Overall target
strength distributions are shown in Figures
5a and 5b
(12K). The
strongest target detected was -30.75 dB at 38
kHz on the EK500, thus suggesting a fish length
of approximately 75 cm. There is a secondary peak at approximately
-40 dB to -45 dB (10-20 cm fish) on the H.A.D.A.S.
and the EK500 (38 kHz).
Catch Data
A
total of four hours of trawling in the scattering
layer yielded 42 Charr of standard lengths between
4.0 cm and 27 cm, plus some Sticklebacks. The hauls are combined in Figure
6a (20K graphs) and show a peak of around
10 cm, doubtless influenced by the selectivity
of the sprat net. Figures
6b, 6c,
and 6d
(7-18K photos) picture the fish of the pelagic. Gill-netting shows the maximum length of
Charr to be approximately 30 cm, and the larger
fish to be Trout, Figure
6d with the largest caught measuring 58
cm (Martin and Shine, 1993).
These larger fish could be expected to evade the
trawl, while smaller ones could escape through
the meshes.
Smaller fish were
not taken until November 1992, when a very large
trawl was undergoing trials from Calanus. Due to the closing of the codend meshes by stretching and further
obstruction by leaves, 62 Charr down to 3.5 cm
were retained, together with similar quantities
of Sticklebacks. The 10 cm peak in the trawl samples would
give target strengths of -48 dB (Lindem, 1984)
or -45.7 dB (Love, 1977), which coincides quite
well with the secondary peak in the target strength
distribution noted above.
Vol
105, The Scottish Naturalist: Pelagic Fish
and Zooplankton in Loch Ness p210
Fish Numbers
and Depth Distributions
The
H.A.D.A.S. runs were made in the South Basin in
August 1988, and showed total numbers of nearly
1,500 fish/ha (Figure
7a, 17K graphs). The mean of the May 1992 EK500 runs in
the North Basin showed a total of 340 fish/ha
(Figure
8a, 14K). Both these estimates include scattering layer
targets. The
remainder amount to no more than 20 fish/ha in
the North Basin (EK500 38 kHz, Figure
8b) and approximately 400 fish/ha in the
South Basin (H.A.D.A.S. 49 kHz (Figure
7b).
The depth distribution
shows all target strength classes to be most numerous
around the area of the scattering layer, but with
substantial numbers of the larger fish above it.
Vertical Migration
The
dusk 5.0 km run with the H.A.D.A.S., and the EK500
midnight runs (Figures
9a and 9b,
12K graphs), clearly show the smaller targets
migrating to the extent that they are almost absent
(too shallow to be surveyed) in the EK500 records.The
larger fish migrate to a much lesser extent; indeed
it would appear that the migration becomes progressively
less with increasing size. It is interesting that, despite the drop
in mean numbers from 341/ha to 31/ha at night,
the EK500 SA figure (total integrated value) has
barely changed, showing the very low 'acoustic
weight' of the scattering layer targets (Figure
10).
North-South
Density Gradient
The
EK 500 length run, made in May 1992 from north
to south along the axis of the loch, showed patchy
numbers but a reasonably consistent target strength
distribution. There was a clear increase in fish numbers
from north to south (Figure
11, 15K and Figure
11a , 25K charts). This confirms previous qualitative observations
throughout the 1980s, particularly in October
1987 (Shine and Martin, 1988).
While this run
was being made, a Clarke-Bumpus zooplankton sampler
was towed at approximately 20 m, close to the
scattering layer. Settled volumes of zooplankton also showed
a definite increase towards the south. A mean of only 8.0 fish/ha was found above
the scattering layer, as opposed to 419 fish/ha
within it. However,
a substantial portion of the biomass (mean SA
2.64) lies above the 20-40 m scattering layer
depths (mean SA 6.39). This again emphasises the low biomass of
the majority of individuals in the scattering
layer.
Overall Estimates
The
patchy distribution makes overall estimates speculative. The mean integration figure apportioned between
the various size classes of the target strength
distribution gives the numbers per hectare. A mean weight can be estimated through
the length/weight charts (Figure
6) by converting the target strength classes
to lengths. The
mean weight is then multiplied by the numbers
given by the integration.
Based upon the
target strength distribution, given by the 10.5
nautical mile runs and the mean of the length
run integration figures, the mean numbers are
427 fish/ha and the biomass 3.1 kg/ha. Thus the resident pelagic fish stock would
be 2,433,900 with a biomass of 17,670 kg. This estimate is exceeded by its statistical
confidence limits, because of the patchiness of
the numerical distribution.
Zooplankton
In
Figures 12a,
12b,
12c,
12d and 12e
(14K graphs) plankton diurnal migration diagrams
are presented, which show that from at least March
onwards a pronounced vertical migration takes
place involving the copepods Diaptomus
gracilis and Cyclops strenuus abyssorum. These are the dominant species. The Cladocera are present in much fewer numbers,
and Bosmina
coregoni in particular appears at the greater
depths (Figure
12b). Daphnia
hyalina appears to migrate to some extent
while Bosmina does not. The larger
predators, such as Leptodora
kindti, Bythotrephes
longimanus and Polyphemus
pediculus, have not been caught in sufficient
numbers to be sure of their movements, although
underwater camera work in 1992 showed diurnal
migration in Leptodora. In Lake Huron (Canada),
however, Bythotrephes
has been reported by Vanderploeg, Liebig and
Omair (1993) to lie in a narrow diurnally-migrating
horizontal band and to favour Cladocera as prey.
In Loch Ness,
Cyclops
has occasionally been observed to be concentrated
at the surface by day, the reverse of the usual
pattern (Figure
12b). Total numbers are seen to increase as the
summer advances. Large variations also occur through horizontal
transport, particularly due to internal seiche
movements, for example in October 1985. Larger numbers have been associated with
denser scattering layers (Marjoram, 1993). Levy, Johnson and Hume (1991) also report changes
in fish distribution due to internal seiche.
By day, total
zooplankton numbers are usually greatest just
above the scattering layer, but sometimes coincide
with it. Migration of the copepod element of the zooplankton
is very similar to that of the scattering layer.
During the length
run (Figure
11, 15K), zooplankton numbers increased
from north to south. All species were more numerous
in the south, although there were differences
in percentage composition. Cyclops increased from 26% to 64% while Diaptomus decreased from 45% to 28%. Bosmina decreased from 8% to 2%.
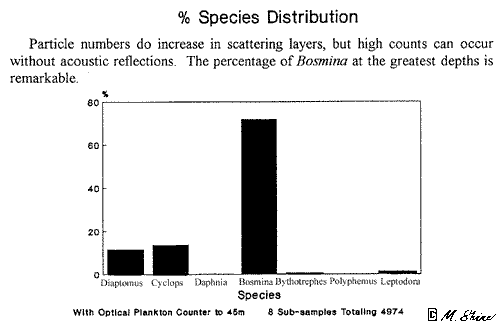
South Basin tows
with the optical plankton counter within deep
acoustic scattering layers (Figure
13a, 46K chart and graph) do show increases
of particle numbers, mostly approximately 0.25
mm, within the layers. However, the greatest peaks in particle
densities can be from depths where there are no
acoustic layers at all. The plankton net attached to the optical
counter yielded the results shown in Figure
13b (7K). The most notable feature is the great preponderance
of Bosmina
at these greater depths.
Scattering
Layer Movements
The
nightly rise appears to be light triggered, commencing
as the light falls to approx 1.0 m/s at 10 m depth
(Marjoram, 1993). The dawn sinking occurs as the light reaches
the same level. It has already been observed that the layer
does not necessarily establish in the thermocline
by day, but very often does (Shine and Martin,
1988). In the summer of 1992 the mixed layer was 12oC
or more, and the scattering layer did establish
in the thermocline at a lower temperature. Experiments with an inverted transducer showed
that the layer does not necessarily rise to the
surface at night but remains at approximately
5.0 m until dawn (Marjoram, 1993).
In the southern
fixed station experiments of 1984 (Shine and Martin,
1988), the scattering layer was seldom entirely
absent, even during north-east flowing internal
seiches. In
the North Basin, however, it is notable that the
scattering layer is strongest during north-east
winds and may disappear entirely during south
westerlies.
Frequently there
is an across-loch tilt in the scattering layer. Figure
14 (29K charts) shows tilting during a
period of south-west winds, with the layer deeper
on the southern shore. Another feature is the presence of 'ramps'
in the scatterers, sloping down towards the wind
direction. They
are approximately 440 m long and slope at an angle
of approximately 4o.
Discussion
The
difference in target strength of the smaller scattering
layer targets recorded by the various systems
is of some interest. To begin with, it would seem that the various
peaks are not the result of optimum detection
thresholds, since total numbers are very similar. In an example on the EK500, 1,600 targets/ha
were recorded at 120 kHz and 2,120 targets/ha
at 38 kHz simultaneously, whereas the target strength
peaks were at -70 dB and -62d B respectively (Mr.
Erik Stenerson, pers. comm.).
Differences could
result from calibration or time-varied gain (T.V.G.),
but results have been comparable on three separate
occasions. It
may be that resonance effects are responsible. There has been little acoustic work on
very small fish.
Burczynski, Michaleitz
and Marrone (1987), in a survey of Rainbow Smelt
Osmerus
mordax, found that Love's (1977) formula appeared
to underestimate the length of 5.6 cm fish by
approximately 2.0 cm. Another point is that very few of the fish
surveyed will be on the beam axis and so be in
true dorsal aspect.
This will reduce
signals even though the beam pattern may have
been compensated. Mr. R. Johnson of Biosonics (pers. comm.)
suggests that the reduction will be greatest at
the higher frequencies, which is consistent with
the pattern of our observations. The low target strengths, especially at
the higher frequencies, have introduced the suggestion
that the individuals of the scattering layer are
too small to be fish. Other candidates are chironomid larvae and
pupae, larger zooplankton such as Leptodora
or perhaps some unrecorded crustacae.
Vol
105, The Scottish Naturalist: Pelagic Fish
and Zooplankton in Loch Ness p227
Shine and Martin
(1988) reported 4th instar chironomid larvae,
particularly Sergentia
sp., taken in the pelagic during plankton
hauls. Larvae
have also been observed in quantity in the stomachs
of pelagic Charr (Martin and Shine, 1993). Chironomid
pupae have been collected by tow-nets within
the scattering layer, particularly in May. It has been observed that, when placed in containers,
the pupae do not necessarily emerge for two or
three days. Therefore it seems possible that they may adopt
a planktonic migration strategy while preparing
for final emergence. Hauls taken later in the year, however, do
not always yield significant numbers of chironomids
even though the scattering layer is strengthening.
Experiments have
been conducted by introducing quantities of 4th
instar chironomid larvae and pupae into an echo-sounder
beam (Lowrance Mach II, 50 kHz) at ranges up to
18 m. Only
occasional traces were obtained, thus suggesting
that detection was dependent on targets close
to the beam axis, perhaps in multiples. Nevertheless, detection can be achieved and
chironomids must be regarded as an important component
of the pelagic community. Work should be undertaken to establish
the target strength of chironomids.
Leptodora
may be dismissed as a candidate, since it is not
present in winter. In March 1991, for example, during a plankton
diurnal, a scattering layer was observed in the
absence of Leptodora (Figure
12a). The optical plankton counter showed
the greatest concentration of smaller particles
in zones not producing acoustic reflections. Therefore zooplankton are unlikely candidates.
With
regard to other species, not hitherto recorded
from Loch Ness, it should be born in mind that
no plankton net, nor any other sampler, in ten
years of work has ever shown traces of any such
organism; nor has any been found in the sediments. No pelagic Trout or Charr examined (Martin
and Shine, 1993) had consumed anything unusual.
The
trawling of 3.5 cm Charr in the pelagic during
November 1992 suggests a very low growth rate. In Martin and Shine (1993) it is shown
that the 1+ Charr are between 4.0 cm and 7.0 cm
long. The new material includes 0+ fish smaller than
this (Mr. R.B. Greer, pers. comm.)
We have observed
spawning Charr in December, which emphasises how
small 0+ fish could be in the earlier summer months. No significant genetic differences have been
observed between Loch Ness pelagic and benthic
Charr (Dr. Sheila Hartley, pers. comm.) and a
fecundity study by Meacham (1993) shows consistent
Vol
105, The Scottish Naturalist: Pelagic Fish
and Zooplankton in Loch Ness p228
egg size increase
from summer to December. It seems unlikely, therefore, that there
is a separate spawning population giving rise
to such small fish in the autumn.
A further point
is that in the earlier months, as in March 1991,
the scattering layer is only detectable on more
powerful echo-sounders. This suggests an increase in individual
target strength (i.e. growth) as the year proceeds.
The scattering
layer diurnal migration is similar to that of
the copepods, but these do not occur significantly
in the fish diet, which is similar for Charr between
4.0 cm and 30 cm. Bythotrephes
and Daphnia are the main prey items, followed
by Leptodora. Only the smallest Charr and some of the Sticklebacks
contained some copepods. It was found by Fryer (1957) that Cyclops
strenuus
abyssorum is predatory upon Diaptomus gracilis. Loch Ness Cyclops have also been observed to feed on animal material, including
Diaptomus
(Mr. K.W. Heath, pers. comm.). A limited amount of plant material is also
consumed, and perhaps this is why Cyclops maxima may sometimes be observed near the surface by day. Diaptomus shows the greatest diurnal migration, and this could be
a response to predation by Cyclops
and other zooplankton rather than by fish. In any case, the scattering layer generally
lies below the copepod maxima. It seems reasonable, by contrast, to suppose
that the much smaller numbers of Daphnia are a reflection of fish predation observed by Martin and
Shine (1993).
At the same time,
the smaller range of Daphnia migration may account for the decrease
in the migration tendencies of the larger Charr. The migration of the scattering layer, if composed
of fish, is perhaps a little surprising if one
of the main food sources is Daphnia,
which does not migrate to the same extent.
It could, however,
be an energetic strategy similar to that exhibited
by the Sculpin Cottus
extensus, a larval fish of North America,
which is a benthic feeder during the day and rises
into the warmer surface waters at night. This speeds digestion and so growth rate (Wayne
and Neverman, 1988). The scattering layer migration could be
a predation response, but none of the pelagic
Charr had consumed fish, and only a few of the
smallest Trout take Charr of <6.0 cm.
The autumn Daphnia
peak noted by Maitland (1981) and ourselves (Figures
12d and 12e)
could perhaps be explained as the larger Charr
move inshore to spawn, thus reducing the predation
pressure. Other
factors could be that the very large water mass
of Loch Ness retains its heat longer than smaller
lakes. Also
there is generally an injection of allochthonous
material (potential food) during equinoctial storms
which bring the rivers into spate.
Vol
105, The Scottish Naturalist: Pelagic Fish
and Zooplankton in Loch Ness p229
The Bosmina
do not seem to migrate at all, and are consumed
by many of the Charr, although seldom in significant
numbers. The
considerable numbers of Bosmina
observed from the thermocline downwards suggests
that they are filter feeding a detrital or microbial
food source. In this connection, the persistent north/south
gradient in fish, and often in zooplankton, is
interesting. This presents something of a paradox, since George and Jones (1987) had found
that both conductivity and chlorophyll-a maxima lie consistently in the North
Basin. They
suggested that this was caused by the slightly
richer northern catchments. An explanation could be that allochthonous organic material brought
in by rivers of the much larger, and wetter, southern
catchments is much more important than the primary
productivity.
If this material
is processed through the microbial element, it
could find its way to the zooplankton, particularly
the filter feeding Cladocera, and so to the fish.
In this respect,
the results of the length run along the axis of
the loch are somewhat surprising, in that the
cladoceran Bosmina decreases in percentage towards
the south. This
species is the one most likely to be utilising
allochthonous inputs, and so might be expected
to form a greater percentage of the plankton composition
in the south. However, it should be borne in mind that the
samples were from a depth of 20 m, and that some
thermocline tilt might have been present due to
a light north-east wind. This could have affected the observed distributions. Numerically all species increased in the South
Basin.
The increase in
fish numbers towards the south is particularly
pronounced in the smaller targets of the scattering
layer. During the 'steady state' produced by the normal
south-west winds, these scatterers would spend
the majority of their time close to the thermocline
and therefore in the south-west return current
produced. By
contrast, a number of length runs have shown that
zooplankton numbers at the near surface increase
down wind (unpublished data).
The
deeper scatterers would only rise into the north-east
surface drift for the six hours of the short northern
nights. Shine and Martin (1988) have shown that the
scattering layer targets move passively in water
currents. Therefore their position in the return current
will tend to increase their density towards the
south. By contrast, during periods of north-east winds,
it is noticeable that the scattering layer beneath
the northern fixed station strengthens appreciably. The other structures noted in the scattering
layer, such as the cross-loch tilts, are consistent
with the circulation of the surface drift due
to Coriolis forces diverting the wind drift to
the right (George, 1981), i.e. to the southern
shore in a south-west wind.
Vol
105, The Scottish Naturalist: Pelagic Fish
and Zooplankton in Loch Ness p232
The 'ramps' are
of interest, and may represent some form of internal
mixing 'fronts' or a reaction to vertical movements
induced by internal waves.
Summary
Hydroacoustic
in situ
target strength assessment techniques have been
applied to the pelagic zone of Loch Ness. The bulk of the targets are distributed
down to the thermocline at densities from 300/ha
to 1,000/ha. Over 80% of these targets are of very small
size, forming a distinct scattering layer. In contrast to the larger fish, this layer
makes nightly migration to the near surface.
The
various hydroacoustic systems give different target
strengths for the small scatterers, between -75
dB and -57 dB, possibly due to a resonance phenomena. The disparity of target strength is much
less for the larger size classes. The identity of individuals comprising the
layer is discussed, and juvenile Charr together
with chironomids are suggested.
Over 80% of the
zooplankton standing crop are copepods which make
pronounced diurnal vertical migrations.
The cladoceran
Daphnia,
which is a main food source for the pelagic Charr,
does not migrate to the same extent, and perhaps
this accounts for the lesser migration shown by
the larger fish.
Bosmina,
a small cladoceran filter feeder, exhibits no
vertical migration, and is often found deep within
the thermocline and below it. It is speculated that the cladocerans,
particularly Bosmina, may utilise allochthonous organic material through the microbial
element of the plankton.
There are consistently
larger numbers of fish in the South Basin, and
greater zooplankton densities are sometimes associated
with this. This
is a paradoxical observation, since George and
Jones (1987) had previously established that conductivity
and chlorophyll-a
concentrations were greater in the north. Again, it is suggested that this is due
to allochthonous inputs.
It has been noted
that the scattering layer can be more dense in
the North Basin with north-east winds, and it
is suggested that this results from the individuals
spending most of the time in the deeper return
current caused by the surface drift. Structures, such as tilts and ramps, in
these scatterers show the close association of
physical events in the vicinity of the thermocline
with the distribution of the scattering layer.
Vol
105, The Scottish Naturalist: Pelagic Fish
and Zooplankton in Loch Ness p233
Acknowledgements
Most of our material
has resulted from the efforts of volunteers aboard
the Loch Ness and Morar Project vessel Ecos,
owned and skippered by Mr. John Minshull. We also acknowledge the support of the
Loch Ness District Salmon Fisheries Board, and
particularly Mr. W. Hastings, the bailiff.
The Simrad company
has loaned most of the acoustic equipment, and
we are also grateful to the crew of this company's
demonstration vessel Simson Echo. Particular help was given by Mr. Barry Pardey,
Mr. Erik Stennerson and Mr. David Wilson. Dr. Torfin Lindem operated the H.A.D.A.S. system.
D.A.F.S. Aberdeen
assisted for some days in August 1988 with their
trawler Goldseeker, and Dr. Richard Ferro has also made
material available from trawling trials aboard
Calanus in 1992.
Invaluable advice
has been given by Dr. J. Simmonds of D.A.F.S.
and Dr. Annie Duncan of Royal Holloway College. Special thanks are due to Mr. R.A. Bremner
of the Official Loch Ness Exhibition Centre for
his continued support and provision of a headquarters
for the Loch Ness and Morar Project.
References
Baker,
P.F. and Westwood, M. (1960). Underwater detective work. Scotsman,
14th September 1960.
Bodholt,
H., Nes, H. and Solli, H. (1988). A new echo-sounder-system for fish abundance
estimation and fishery research. In: Proceedings of the International Council for the Exploration of the Sea. C.M.
1988/B:11. Session P. [Aberdeen: Simrad Company].
Burczynski,
J.J., Michaleitz, P.H. and Marrone, G.M. (1987). Hydroacoustic assessment of the abundance
and distribution of Rainbow Smelt in Lake Oahe. North
American Journal of Fisheries Management, 7: 106-116.
Craig,
R.E. and Forbes, S.T. (1969). Design of a sonar for fish counting. Fiskeridirektoratets
Skrifter, 15: 210-219.
Vol
105, The Scottish Naturalist: Pelagic Fish
and Zooplankton in Loch Ness p234
Ehrenberg,
J.E. (1978). The dual-beam system, - a technique for making
in situ measurements
of the target strengths of fish. In: Proceedings of the Acoustics in Fisheries Conference. Faculty of Maritime
and Engineering Studies, Hull College of Higher Education, 26-27
September 1978. University of Bath, England.
Ehrenberg,
J.E. (1979). A comparative analysis of in
situ methods for directly measuring
the acoustic target strength of individual fish. Journal of Oceanographic Engineering, 4: 141-152.
Fryer,
G. (1957). The
food of some freshwater cyclopoid copepods and
its ecological significance. Journal of Animal Ecology,
26: 263-286.
George,
D.G. (1981). Wind-induced
water movements in the south basin of Windermere. Freshwater
Biology, 11: 37-60.
George,
D.G. and Jones, D.H. (1987). Catchment effects on the horizontal distribution
of phytoplankton in five of Scotland's largest
freshwater lochs. Journal of Ecology, 75: 43-59.
Hartman,
J., Dahm, E., Dawson, J., Doering, P., Jurgensen,
S., Lindem, T., Loffler, H. and Raemhild, G. (1984). European Inland Fisheries Advisory Commission
(EIFAC). Experiments on Pelagic Fish Stock Assessment
by Acoustic Methods in Lake Tegel. EIFAC Occasional Paper No. 17: 1-26. Rome: Food and Agricultural
Organisation of the United Nations.
Jurvelius, J., Lindem, T. and Louhimo, J. (1984). The
number of pelagic fish in lake Paasivesi, Finland,
monitored by hydroacoustic methods. Fisheries Research, 2: 273-283.
Kubecka,
J., Duncan, A. and Butterworth, A.J. (1993). Large and small organisms detected in the
open waters of Loch Ness by dual-beam acoustics. Scottish
Naturalist, 105: 175-193.
Levy,
D.A., Johnson, R.L. and Hume, J.M. (1991). Shifts in fish vertical distribution in
response to an internal seiche in a stratified
lake. Limnology
and Oceanography, 36: 187-192.
Lindem,
T. (1984). European
Inland Fisheries Advisory Commission (EIFAC). Inter-calibration
Exercise, Lake Constance, 23-29 October 1983.
Hydroacoustic Data Report. Geneva: Food and Agriculture Organisation of the United
Nations.
Love,
R.H. (1977). Target
strength of an individual fish at any aspect. Journal of the Acoustical Society of America,
62: 1397-1403.
Maitland,
P.S. (Ed.) (1981). The Ecology of Scotland's Largest Lochs: Lomond, Awe, Ness, Morar and Shiel. Monographiae Biologicae, Vol. 44. The Hague, Junk.
Marjoram,
R.S. (1993). An
Investigation of the Identification and Behaviour
of an Acoustic Scattering Layer in Loch Ness,
Scotland. B.Sc. Dissertation, Worcester College of
Higher Education.
Martin,
D.S. and Shine, A.J. 1993). The food and feeding relationships of pelagic
fish in Loch Ness. Scottish Naturalist, 105:
149-174.
Vol
105, The Scottish Naturalist: Pelagic Fish
and Zooplankton in Loch Ness p235
Meacham,
N. J. (1993). The Fecundity and Associated Ecological Factors
of the Arctic Charr,
Salvelinus alpinus, and Brown Trout, Salmo
trutta, in Loch Ness, Scotland. B.Sc. Dissertation. University of Hull.
Shine,
A.J. and Martin, D.S. (1988). Loch Ness habitats observed by sonar and underwater
television. Scottish
Naturalist,
100: 111-199.
Traynor,
J.J. and Ehrenberg, J.E. (1979). Evaluation of the dual-beam acoustic fish target strength measurement
method. Journal of the Fisheries Research Board of
Canada, 36: 1065-1071.
Vanderploeg,
H.A., Liebig, J.R. and Omair, M. (1993). Bythotrephes predation on Great Lakes' zooplankton
measured by an in situ method: Implications for zooplankton community structure. Verhandlungen der Internationalen Vereinigung fur Theoretische und Angewandte
Limnologie, 25: 349.
Wayne,
A.W. and Neverman, D. (1988). Post-feeding
thermotaxis and daily vertical migration in a
larval fish. Nature, 333: 846-848.
Received
June 1993
Mr. Adrian J. Shine, Loch Ness and Morar Project,
Loch Ness Centre, DRUMNADROCHIT, Inverness-shire IV3 6TU.
Mr. David S. Martin, Loch Ness
and Morar Project,
Loch Ness Centre, DRUMNADROCHIT, Inverness-shire IV3 6TU.
Miss Rosalind S. Marjoram, Loch Ness and Morar Project,
Loch Ness Centre, DRUMNADROCHIT, Inverness-shire IV3 6TU.

{kind=link}