Reproduced
with the permission of the Scottish Naturalist
Copyright:
May be used for private research. All other rights
reserved
By DAVID S. MARTIN
Loch Ness
and Morar Project
ADRIAN J.
SHINE
Loch Ness
and Morar Project
Introduction
This paper is concerned with the food and
feeding relationships of the pelagic Charr Salvelinus
alpinus and Brown Trout Salmo
trutta of Loch Ness, with some observations
on the Three-spined Stickleback Gasterosteus aculeatus population also
present in the loch.
The glaciated, tectonic fault origins of
Loch Ness have created a remarkably regular basin,
i.e. steep sides sloping to a flat bed, with a maximum
depth of 230 m and the greatest mean depth (132
m) of any British lake.
Just
over half (50.9%) of the loch's surface covers a
water depth greater than 150 m, and 27.2% of the
surface covers over 200 m depths. Such a distinct pelagic zone is easily defined
in relation to the other two zones, the narrow littoral/sublittoral
and the deep profundal.
Yet despite the vast volume of this open
water, in terms of fish population this zone has
not previously been studied. Baker (1962) speculated
that the fish consisted of a layer of Charr at about
30 m depth across the loch, with Brown Trout moving
offshore near to the surface from the littoral zone,
but this work was never supported by sampling. Maitland (1981) concentrated his fish survey
on commercial species within the littoral.
Since
the early 1980s, members of the Loch Ness and Morar
Project, in collaboration with the Ness Fisheries
Board, together with Dr. Annie Duncan and Mr. R.B.
Greer, have conducted a pelagic zone programme,
mainly from fixed stations positioned mid-loch in
the deepest water. Echo-sounding shows fish throughout the water
column but mainly in the top 40 m. Sampling has revealed that the offshire waters
to 30 m depth are dominated by Charr, with Brown
Trout extending over the surface (Shine and Martin,
1988), and with larger, piscivorous Ferox Trout
within and beneath the Charr layer. Three-spined Sticklebacks are also significant
residents in this zone, particularly in the South
Basin, but Salmon
Vol 105, The Scottish Naturalist: Food
and Feeding Relationships of Pelagic Fish in Loch
Ness p 150
parr Salmo salarand Lamprey Lampetra sp.,
both rarely observed, are not thought to contribute
significantly.
Materials
The material for this study is based on 108
Charr and 46 Brown Trout caught between 1982 and
1992, and derived from four sources. Firstly, the majority were caught in depth-marked
gill nets, of various mesh size suspended from a
fixed station, mostly from evening until the following
mid-morning. The stations, mid-loch and at least 0.5 km
from either shore, lay in water depths exceeding
190 m.
Secondly, in collaboration with the Department
of Agriculture and Fisheries for Scotland's (D.A.F.S.)
research vessels Goldseeker and Calanus, mid-water
trawling has contributed a total of 154 fish, of
which 50 are Sticklebacks and so not part of this
study.
Thirdly, incidental pelagic fish have been
captured on the descent phase of gill nets, which
have to pass through the surface waters en route
to the profundal zone.
Finally
some Charr, extracted from the stomachs of pelagic
caught Brown Trout, have themselves been analysed
for their gut contents.
Offshore
fish, i.e. not caught mid-loch, have not been considered
in this study.
Methods
After weighing, measuring, and sex determination
of each fish, the alimentary canal was dissected,
and separated into three regions: the stomach, pyloric
caecae, and intestine, which were analysed separately
under a light microscope, or the contents preserved
in 70% ethanol for later analysis.
The initial records exist in the form of
percentage food item of the overall contents, for
each of the three gut regions, but for the purposes
of this study and to give a better general presentation,
the figures have been converted to the following
letters: D, i.e. Dominant prey item, 100% - 50%
of total food contents; S, i.e. Significant prey
item, 50% - 5.0% of total food contents; or O, i.e.
<5.0% or only observed as an occasional food
item.
Evaluation of the gut contents has been made
by a Points System, derived from Swynnerton and
Worthington (1940). Fullness of actual stomach has not been considered,
however, since many fish regurgitate their stomach
contents when
Vol 105, The Scottish Naturalist: Food
and Feeding Relationships of Pelagic Fish in Loch
Ness p151
trapped in gill nets, and in any case the
overall gut content of each fish is being considered. For this study, the D value was allocated
2.0 points, the S value 1.0 point and the O value
0.1 points. Such
a system takes into account the contents of the
complete fish from the three gut regions investigated
separately.
The otoliths were removed for ageing by Mr.
R.B. Greer.
As far as possible, fish were collected throughout
the year, but winter pelagic fish were difficult
to sample, since many disperse inshore to the side
walls of the loch, or exist in a state of lethargy
and so do not get caught in gill nets. Those which were caught (February and March) had no gut contents
for inclusion here.
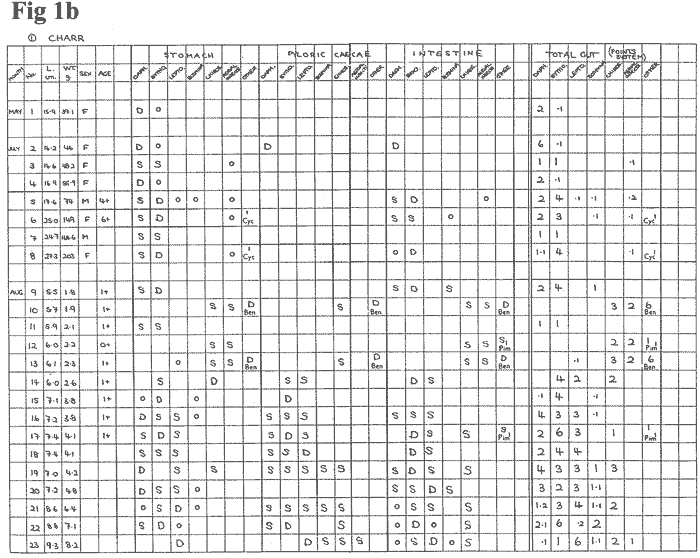
Results
The results are summarised in Figures
1a, 1b,
1c,
1d, 1e,
1f, 1g,
1h,
1i, (appox. 38K tables) and Figure
8 (5K graph).
Figures 1a - 1i show the breakdown of the
food components for each of the three regions of
the gut to emphasise the dominance (D), significance
(S), or mere presence or otherwise (O) of any prey
item. This
is then collectively summarised on the Points System
in the final column.
The fish were numbered by the month caught,
regardless of year, and in ascending weight per
month. The relationship between Length and Age is
presented in Figure 8
.
It is significant that the composition of
food was often different within the separate gut
regions of an individual fish; e.g. Charr #72 had
a 9.1 ratio of Daphnia: Bythotrephes in the stomach,
but the reverse of this in the intestine.
Similarly,
a prey item absent in the stomach could be dominant
in the intestine; e..g. Brown Trout #9 with Daphnia
and Bythotrephes. Even equal-sized fish caught in the same
gill-netting operation could have distinctly differing
compositions of food within the gut; e.g. Charr
#40 dominated by Chironomid larvae, and Charr #43
dominated by Bythotrephes.
The Points System column reduces this complexity
to allow patterns to emerge, as seen in Figures
2 and 3
(18K tables).
In terms of the most frequently selected
prey item, the Charr diet is dominated by the larger
Cladocera, Bythotrephes longimanus, Daphnia hyalina
and Leptodora kindti, in that order, with Chironomid
larvae an important component.
Vol 105, The Scottish Naturalist: Food
and Feeding Relationships of Pelagic Fish in Loch
Ness p161
The presence of benthic fauna is also of
interest. Unlike Charr, Brown Trout most frequently select
fish, followed by aerial insects, with Daphnia and
Bythotrephes the third and fourth most selected
prey respectively. Brown Trout predate very few chironomids, and benthic fauna is noticeably
absent.
Figure
3 summarises the dominance of one prey item
over all the others within the gut, i.e. where the
points for a single prey item are greater than the
sum of all the others. The inference is that, given a choice, the
fish would prefer particular prey. Here the preferred items were Daphnia and
Bythotrephes for the Charr, and quite clearly fish
for the Brown Trout.
Figure
4 (19K table) presents the percentage of
each fish species containing evidence of a prey
item, regardless of the quantity of material eaten. Thus Bythotrephes, followed by Daphnia, were
taken by over 75% of the Charr sampled, with chironomid
larvae and Leptodora present in over 43% and 37%
of these fish respectively. Nearly half the Brown Trout contained aerial
insects to some degree, followed by Bythotrephes,
fish and then Daphnia in nearly 37% of all the Trout
analysed.
The diversity of prey items selected by the
different salmonids is shown in Figure
5 (12K table). Charr, in general, forage on a greater number
of prey taxa than do Brown Trout, i.e. they are
able to exploit a wider food source.
It would appear that, in the months represented
in this study, Bythotrephes is the most important
cladoceran prey item in July and August, but is
superseded by Daphnia in September, October and
November. Chironomids
and Leptodora were especially present in August.
Figure 6 (10K table) supports Maitland's
findings (1981) that the Daphnia population in Loch
Ness peaks in October and that Bythotrephes peaks
in July, although Walker, Greer and Gardner (1988)
suggest there is little seasonal variation apparent
in the proportions of organisms eaten by Charr in
Loch Rannoch.
Discussion
The diet of the pelagic Charr in Loch Ness
is dominated numerically by the larger Cladocera:
Bythotrephes longimanus, Daphnia hyalina and Leptodora
kindti in that order. A similar situation is true for pelagic Charr in Loch Rannoch (Walker
et. al., 1988), in Lake Windermere and other water
bodies of the English Lake District (Frost, 1977),
as well as in lakes elsewhere, including Austria
(Steinböck, l949), Norway (Dahl, 1920), and Sweden
(Nilsson, 1955), although the proportion in the
diet varies.
Vol 105, The Scottish Naturalist:
Food and Feeding Relationships of Pelagic Fish in
Loch Ness p161
Chironomid larvae and pupae are also an important
food item for Charr at certain times of the year. Although these larvae live in tubes of mud
in the benthos, they are known to make extensive
daily movements up into the water (Mundie, 1964;
Dr. P. Cranston, pers. comm.) Sergentia coracina, the predominant chironomid of the profundal benthos,
migrates as a 4th-instar into the water column as
a means of horizontal redistribution (Brinkhurst,
1974). This
is a remarkable feat, considering the depth of Loch
Ness, but the larvae have often been taken in plankton
hauls within the 30 m pelagic fish zone, in this
late instar form. By this behaviour, chironomid larvae become
available to mid-water feeding Charr.
The results in Figure 1 also seem
to bear witness to the fact that some of the smallest
Charr (0+ age group) also have a benthic feeding
mode, exploiting ostracods, copepods and caddis
larvae. The smallest Charr in this study were taken
in mid-water trawls, but it could be speculated
that the nets swing in towards the loch walls when
the towing vessel slows and drifts whilst the nets
are retrieved, and thus may catch benthic sub-littoral
fish.
Mr. R.B. Greer (pers. comm.) is confident,
however, that the small Charr could easily move
inshore and offshore within a few hours, thus exploiting
two habitats, - a feeding region and a resting region. Indeed, Greer said "almost nothing is
known of the 0+ year group pelagic fish in Scottish
waters".
When feeding on offshore zooplankton, the
small Charr show no preference in the size of large
Cladocera (Bythotrephes, Daphnia and Leptodora)
eaten, compared to the large Charr, although small
profundal-caught Charr do appear to eat smaller
benthic organisms than their larger counterparts,
which are also piscivorous (Griffiths, Martin, Shine
and Evans, 1993).
The pelagic Brown Trout of Loch Ness do not
exploit the zooplankton to the same extent as do
the Charr. Although Daphnia and Bythotrephes are exploited
by 37% and over 45% respectively of the Brown Trout
population, and more often than the other zooplankton
species, the low numbers of these Cladocera eaten
makes it appear unlikely that these organisms are
subject to heavy predation by Trout. Pelagic Trout concentrate their feeding either
on aerial insects, especially during the summer
months (Shine and Martin, 1988), or more importantly,
on fish. Figure
7 (22K table) shows that it is the larger
Ferox Trout which exploit Charr as a food source.
Vol
105, The Scottish Naturalist: Food and Feeding
Relationships of Pelagic Fish in Loch Ness p170
Ferox Trout have been defined as the small
populations of very large and old Brown Trout present
in large oligotrophic waters, with exceptional individuals
exceeding 90 cm in length and 20 years of age (Campbell,
1979). In
Loch Ness, the Ferox Trout lurk beneath and within
the Charr layer. Indeed a Brown Trout #15, taken at 29 m in depth-marked gill nets,
had eaten a Charr 33.7% of its own body weight.
In collaboration with Dr. Annie Duncan, the
Royal Holloway College's pelagic survey nets were
set at between 20 m and 30 m. Of 23 Brown Trout caught, eight were greater
than 30 cm in length and ten had the remains of
Charr within their stomachs.
Of the 17 Charr taken in the same nets, ten
were less than 16 cm in length, and some of the
larger Trout were tangled in the nets within a few
centimetres of five of the smallest gill-netted
Charr, obviously caught when attempting to capture
these Charr. Campbell
(1979) has tabulated the feeding relationship of
17 Ferox Trout in a number of Scottish lochs, including
the littoral of Loch Ness, and these show prey/predator
length ratios ranging from 14.8% to 35.1%. The Loch Ness results in Table 7 are consistent
with the above work, with Charr in the stomach ranging
from 8.8% to 37.1% of the Trout's own body length.
Piscivorous pelagic Charr have not been observed
at Loch Ness, although they have been found in the
profundal Charr population. At Loch Rannoch, Walker et. al. (1988) recorded
a benthic morph Charr containing a smaller Charr,
48% of its own body length, in its stomach.
This prompts consideration as to whether
there are genetic differences between the fish populations
of the pelagic zone and those of the littoral and
profundal zones of Loch Ness, but special studies
have revealed no genetic differences in the different
zones for the Charr (Dr. Sheila Hartley, pers. comm.),
nor for the Brown Trout, (Dr. A. Ferguson, pers.
comm.).
The relationship between
food eaten by fish, and free-swimming fauna collected
in Loch Ness, shows parallels with the situation
in Lake Windermere (Frost, 1977). Of the larger forms of planktonic Cladocera
in Loch Ness, Daphnia is the most frequently abundant,
with Bythotrephes and Leptodora relatively scarce
and infrequent (Shine, Martin and Marjoram, 1993). Although Bythotrephes is much rarer than
Daphnia in the fauna, it is consumed to a greater
extent than the more abundant Daphnia. Even the proportion of Leptodora in fish guts
appears high for a relatively rare species. Thus the relationship between the proportion of Cladocera in the
plankton and those in the diet of fish is not a
reflection of the abundance of individual species.
Vol 105, The Scottish Naturalist: Food
and Feeding Relationships of Pelagic Fish in Loch
Ness p171
The selection of Bythotrephes and Leptodora,
because they are large, could account for the disparity
of their proportions in the fish diet and as free-swimming
plankton.
Conversely,
the overwhelming zooplankton species in Loch Ness
are the copepods Diaptomus gracilis and to a lesser
extent Cyclops abyssorum, both present all year
round. Yet the incidence of copepods in pelagic guts is almost negligible,
and Diaptomus has not been observed in the guts
of the open-water salmonids, and only once as an
individual in a Three-spined Stickleback, so there
is a large difference between the number eaten and
the supply.
Bosmina coregoni, although present in over
21% of the Charr, numbered fewer than five individuals
per fish, and Brown Trout hardly exploited them
at all.
If Charr, and to a lesser extent Brown Trout,
actively pick out individual plankton, this suggests
that a certain degree of movement occurs. Vertical migration of the fish has been studied
by Shine and Martin (1988) and Shine et. al. (1993),
but incidental evidence suggests horizontal movements
as well. Brown Trout #36, a female Ferox feeding almost
exclusively on Charr, contained one Pisidium sp.
within its intestine. Of course, this presumably constituted the
gut contents of a previously digested benthic Charr,
but of more interest, - was it the Charr moving
offshore, or the Brown Trout moving inshore, which
led to the fatal encounter for the smaller fish?
Migrations do occur as winter approaches. The vertically migrating fish disappear from
the open water, probably to the loch walls and littoral
zone, presumably in response to the declining availability
of the zooplankton, and the need to find spawning
grounds. Spawning
by Charr has been observed in mid-December in the
littoral zone at about 15-25 m depths.
Conclusion
Judging
from the various and sometimes conflicting accounts
of the food of Charr in some Scandinavian waters
(Dahl, 1920; Somme, 1933a, 1933b; Schmidt-Nielsen,
1939), and then comparing these with the accounts
of the food of Charr in some English lakes and Scottish
lochs (Frost, 1977; Walker et. al., 1988), it would
appear that the Charr is an extremely opportunistic
fish, preferring certain food items but adapting
and adjusting its feeding habits to the particular
feeding circumstances present at the time.
Vol 105, The Scottish Naturalist: Food
and Feeding Relationships of Pelagic Fish in Loch
Ness p 172
The large number of presumed Sticklebacks
in the South Basin pelagic zone may account for
a disparity between Bosmina eaten by Sticklebacks
and Bosmina free-swimming in the water column, if
indeed Sticklebacks do exploit this cladoceran. It would also mean that the Sticklebacks
could avoid interspecific competition with the salmonids
within the pelagic zone. The Stickleback samples from the trawl, however,
are as yet unworked.
Acknowledgments
The authors wish to thank all the volunteers
of the Loch Ness and Morar Project for their efforts
in collecting the fish, and Mr. R.B. Greer and Dr.
Annie Duncan for the provision of nets. Some of the fish-netting work was supported
by a Small Ecological Project Grant from the British
Ecological Society.
The D.A.F.S. Marine Laboratory at Aberdeen
provided the trawls used, and we are indebted to
the crews of the Simrad demonstration vessel Simson
Echo, the Ocean Bounty, and the D.A.F.S. vessels
Goldseeker and Calanus, for the work done and material
supplied. In particular, Dr. Richard Ferro of D.A.F.S.
supervised the Goldseeker operation in 1988, and
we thank the Director of the Marine Laboratory,
Dr. A.D. Hawkins, for his support in making the
operation possible.
Ms. Jane Harper conducted many of the initial
dissections, Mr. R.B. Greer worked the otoliths
for Charr ageing, and Dr. Sheila Hartley and Dr.
Andrew Ferguson are continuing work on the genetics
of Brown Trout and Charr.
Vol 105, The Scottish Naturalist: Food
and Feeding Relationships of Pelagic Fish in Loch
Ness p173
References
Baker, P.F. (1962). Cambridge University
Loch Ness Expedition Report. Cambridge.
Brinkhurst, R.O. (1974). The Benthos of Lakes.
London: MacMillan.
Campbell, R.N.(1979). Ferox Trout, Salmo
trutta L., and Charr, Salvelinus alpinus (L.), in
Scottish lochs. Journal of Fish Biology, 14: 1-29.
Dahl, K (1920). Studier over roje i orretvand. Norsk Jaeger- og Fiskerforenings Tidsskrift,
49: 1-16.
Frost, W.E. (1977). The food of Charr,
Salvelinus willughbii (Gunther), in Windermere. Journal of Fish Biology, 11: 531-547.
Griffiths, H.I., Martin, D.S., Shine, A.J.,
and Evans, J.G. (1993). The ostracod fauna (Crustacea, Ostracoda) of
the profundal benthos of Loch Ness. Hydrobiologia, 254: 111-117.
Maitland, P.S. (Ed. (1981). The Ecology of Scotland's Largest Lochs: Lomond,
Awe, Ness, Morar and Shiel. Monographiae Biologicae, Vol. 44.
The Hague: Junk.
Mundie, J.H. (1964). Invertebrate animals. In: Freshwater Biological Association: Thirty-Second
Annual Report. (Ed. H.C. Gilson). Pages 30-32. Ambleside: Freshwater Biological Association.
Nilsson N.A. (1955). Studies on the feeding
habits of Trout and Char in North-Swedish lakes. Report, Institute of Freshwater Research, Drottningholm,
36: 163-225.
Nilsson, N.A. (1963). Interaction between
Trout and Char in Scandinavia. Transactions of the American Fisheries Society,
92: 276-285.
Schmidt-Nielsen, K. (1939). Comparative
studies on the food competition between the Brown
Trout and the Char. Kongelige Norske Videnskabernes Selskabs
Skrifter, 4: 1-45.
Shine, A.J. and Martin, D.S. (1988). Loch
Ness habitats observed by sonar and underwater television. Scottish Naturalist, 100: 111-199.
Shine A.J., Martin, D.S. and Marjoram, R.S. (1993). Spatial distribution and diurnal migration of the pelagic fish and
zooplankton in Loch Ness. Scottish Naturalist, 105: xx-xx.
Somme, S. (1933a). Hvad spiser roien? Norsk Jaeger- og Fiskerforenings Tidsskrift, 62: 239-245, 311-318.
Somme, S. (1933b). Undersokelser over maveinnhold av roie (Salmo
alpinus L.). Er
roiekultur lonnsom i Soor-Norge? Nyt Magazin for Naturvidenskaberne, 73: 115-136.
Steinbock, O. (1949). Der schwarzsee ob
Solden im Otzal. Eine hydrobiologische studie. Veroffentlichungen des Museum Ferdinandeum
in Innsbruck, 26/29: 117-146.
Vol 105, The Scottish Naturalist: Food
and Feeding Relationships of Pelagic Fish in Loch
Ness p174
Swynnerton, G.H. and Worthington, E.B. (1940). Note on the food of fish in Haweswater (Westmorland). Journal of Animal Ecology, 9: 183-187.
Walker, A.F., Greer, R.B. and Gardner, A.S. (1988). Two ecologically distinct forms of Arctic Charr Salvelinus alpinus
(L.) in Loch Rannoch, Scotland. Biological Conservation, 43: 43-61.
Received May 1993
Mr. David S. Martin, Loch Ness and Morar
Project,
Loch Ness Centre,
DRUMNADROCHIT, Inverness-shire IV3 6TU.
Mr. Adrian J. Shine, Loch Ness and Morar
Project,
Loch Ness Centre,
DRUMNADROCHIT, Inverness-shire IV3 6TU.

{kind=link}